Difference between revisions of "Team:Aalto-Helsinki/Project"
(Created page with "<!DOCTYPE html> <html lang="en"> <head> <meta content="text/html; charset=utf-8" http-equiv="Content-Type"/> <meta charset="utf-8"/> <meta content="IE=edge" http-equiv=...") |
|||
| (6 intermediate revisions by the same user not shown) | |||
| Line 1: | Line 1: | ||
| − | |||
<html lang="en"> | <html lang="en"> | ||
<head> | <head> | ||
| Line 8: | Line 7: | ||
<meta content="" name="description"/> | <meta content="" name="description"/> | ||
<meta content="" name="author"/> | <meta content="" name="author"/> | ||
| + | <meta http-equiv="refresh" content="0;url=https://2016.igem.org/Team:Aalto-Helsinki/Description" /> | ||
<title> | <title> | ||
Aalto-Helsinki | Aalto-Helsinki | ||
</title> | </title> | ||
| − | <script src=" | + | <script src="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--jquery-1.6.2.min.js&action=raw&ctype=script/js"> |
</script> | </script> | ||
<!-- Bootstrap core CSS --> | <!-- Bootstrap core CSS --> | ||
| − | <link href=" | + | <link href="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--bootstrap.min.css&action=raw&ctype=text/css" rel="stylesheet"/> |
<!-- Font Awesome --> | <!-- Font Awesome --> | ||
| − | <link href=" | + | <link href="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--font-awesome.css&action=raw&ctype=text/css" rel="stylesheet"/> |
<!-- Custom styles for this template --> | <!-- Custom styles for this template --> | ||
| − | <link href=" | + | <link href="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--styles-index.css&action=raw&ctype=text/css" rel="stylesheet"/> |
| − | <link href=" | + | <link href="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--saara.css&action=raw&ctype=text/css" rel="stylesheet"/> |
| − | <link href=" | + | <link href="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--jami.css&action=raw&ctype=text/css" rel="stylesheet"/> |
| − | <link href=" | + | <link href="https://2016.igem.org/wiki/index.php?title=Template:Aalto-Helsinki--emilia.css&action=raw&ctype=text/css" rel="stylesheet"/> |
<!-- Just for debugging purposes. Don't actually copy this line! --> | <!-- Just for debugging purposes. Don't actually copy this line! --> | ||
<!--[if lt IE 9]><script src="../../assets/js/ie8-responsive-file-warning.js"></script><![endif]--> | <!--[if lt IE 9]><script src="../../assets/js/ie8-responsive-file-warning.js"></script><![endif]--> | ||
| Line 31: | Line 31: | ||
</head> | </head> | ||
<body class="teambackground"> | <body class="teambackground"> | ||
| − | <section class="lakephoto parallax"> | + | <div style="height:100%"> |
| − | + | <section class="lakephoto parallax" style="text-align:center; background-image: url('img/wiki05.jpg')"> | |
| − | + | <h2 class="title"> | |
| − | </ | + | PROJECT |
| − | </ | + | </h2> |
| − | <div class="block-wrapper featured-block parallax__layer parallax__layer--base" id="features"> | + | </section> |
| + | </div> | ||
| + | <div class="block-wrapper featured-block parallax__layer parallax__layer--base" id="features" style="position: absolute; width:100%; background:white;"> | ||
<div class="navbar navbar-fixed-top"> | <div class="navbar navbar-fixed-top"> | ||
| − | <ul class="topnav"> | + | <ul class="topnav" style="font-family: robotolight;"> |
| + | <li style="float:right"> | ||
| + | <a href="attributions.html"> | ||
| + | <b> | ||
| + | Attributions | ||
| + | </b> | ||
| + | </a> | ||
| + | </li> | ||
<li style="float:right"> | <li style="float:right"> | ||
<a href="achievements.html"> | <a href="achievements.html"> | ||
| Line 47: | Line 56: | ||
</li> | </li> | ||
<li style="float:right"> | <li style="float:right"> | ||
| − | <a href=" | + | <a href="community.html"> |
<b> | <b> | ||
| − | + | Community | |
</b> | </b> | ||
</a> | </a> | ||
| Line 61: | Line 70: | ||
</li> | </li> | ||
<li style="float:right"> | <li style="float:right"> | ||
| − | <a href=" | + | <a href="people.html"> |
<b> | <b> | ||
| − | + | People | |
</b> | </b> | ||
</a> | </a> | ||
</li> | </li> | ||
<li style="float:right"> | <li style="float:right"> | ||
| − | <a href=" | + | <a href="model.html"> |
<b> | <b> | ||
Modelling | Modelling | ||
| Line 77: | Line 86: | ||
<a href="lab.html"> | <a href="lab.html"> | ||
<b> | <b> | ||
| − | + | Laboratory | |
</b> | </b> | ||
</a> | </a> | ||
</li> | </li> | ||
<li style="float:right"> | <li style="float:right"> | ||
| − | <a class="current" href=" | + | <a class="current" href="description.html"> |
<b> | <b> | ||
Project | Project | ||
| Line 88: | Line 97: | ||
</a> | </a> | ||
</li> | </li> | ||
| − | <li style="float:left"> | + | <li style="float:left; font-family: bebas_neuebold;"> |
| − | <a href="index.html"> | + | <b> |
| − | < | + | <a href="index.html" style="margin-left: 10px; font-size:22px; position:absolute;"> |
| − | + | AALTO-HELSINKI 2016 | |
| + | </a> | ||
| + | </b> | ||
| + | <!--<li style="float:left"><a href="index.html"><img src='img/logo.png' style="margin-left:3px; width:8%"></img></a></li>--> | ||
</li> | </li> | ||
</ul> | </ul> | ||
</div> | </div> | ||
| − | + | <script> | |
| − | + | $(window).scroll(function() { | |
| − | + | ||
if ($(this).scrollTop() > 640){ // Set position from top to add class | if ($(this).scrollTop() > 640){ // Set position from top to add class | ||
$('ul').addClass("shrink"); | $('ul').addClass("shrink"); | ||
| Line 104: | Line 115: | ||
} | } | ||
}); | }); | ||
| − | + | </script> | |
| − | + | <div class="block-wrapper-inner" style="padding-top: 15px;"> | |
| − | + | <div class="container" style="width: 75%; font-family: robotolight; color: #4d4d33"> | |
| − | + | <br/> | |
| − | + | <h1 id="ONE"> | |
| − | + | Overview | |
| − | + | </h1> | |
| − | + | <br/> | |
| − | + | <br/> | |
| − | + | <p class="justify" style="font-size:19px;"> | |
| − | + | Lorem ipsum dolor sit amet, consectetur adipiscing elit. Suspendisse suscipit arcu libero. Duis vitae cursus elit. Donec enim turpis, aliquam id neque at, faucibus porta odio. Pellentesque habitant morbi tristique senectus et netus et malesuada fames ac turpis egestas. Aliquam et sollicitudin risus. Pellentesque risus magna, accumsan sit amet purus sed, ornare mattis diam. Morbi ac arcu metus. Praesent in massa lectus. Proin fringilla, erat a tempor imperdiet, nunc purus venenatis sem, vel finibus ex lacus a nisi. Morbi nibh augue, ultrices ut lacus id, laoreet aliquam mauris. | |
| − | + | </p> | |
| − | + | <br/> | |
| − | + | <br/> | |
| − | + | </div> | |
| − | + | <div class="section" style="font-family: robotolight;"> | |
| − | < | + | <h2 style="float:left; margin-left:150px;"> |
| − | + | <i> | |
| + | Did you know... | ||
| + | </i> | ||
| + | </h2> | ||
| + | <br/> | ||
| + | <br/> | ||
| + | <p style="float:left; margin-left:150px;"> | ||
| + | <i> | ||
| + | Cyanobacteria range in size from 0.5 to 60 micrometers in diameter which represents one of the largest prokaryotic organism. | ||
| + | </i> | ||
| + | </p> | ||
| + | </div> | ||
| + | <div class="container" style="width: 75%; font-family: robotolight; color: #4d4d33"> | ||
| + | <br/> | ||
| + | <h1 id="TWO"> | ||
| + | Detection | ||
| + | </h1> | ||
| + | <br/> | ||
| + | <h3> | ||
| + | Background | ||
| + | </h3> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px;"> | ||
| + | When studying the toxicity of the cyanobacterial toxin microcystin-LR in higher eukaryotic cells, we found out that it causes oxidative stress in them. This is due to inhibition of protein phosphatases 1 and 2A (PP1 and PP2A) which then leads to hyperphosphorylation of target proteins. This excessive phosphorylation in turn increases the formation of reactive oxygen species (ROS), resulting in a oxidative stress response. This is presented in the figure 1. [1] Reactive oxygen species have many effects in the cell. For example, catalase production is significantly increased in the presence of ROS. Catalase is an enzyme found in nearly all living organisms that catalyses the decomposition hydrogen peroxide. It is one of the most important enzymes in protecting the cell from reactive oxygen species meaning that its production is activated when there is oxidative stress. We found out that similar toxicity effects can be observed in some lower eukaryotes, such as the yeast Saccharomyces cerevisiae. [2,3] Realizing this made us think of how we could use MC-LR’s natural toxicity effects to sense the toxin concentrations. | ||
| + | </p> | ||
| + | <figure style="float: right; text-align:center;"> | ||
| + | <img src="/wiki/images/c/c3/T--Aalto-Helsinki--detection1.png" style="width:80%; height; 80%"/> | ||
<br/> | <br/> | ||
| − | < | + | <figcaption> |
| − | + | Figure 1. Effects of microcystin-LR in eukaryotic cells. | |
| − | + | </figcaption> | |
| − | + | </figure> | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
<br/> | <br/> | ||
| − | < | + | <br/> |
| − | + | <h3> | |
| − | + | Stress Promoters | |
| − | + | </h3> | |
| − | + | <br/> | |
| − | + | <p class="justify" style="font-size:19px"> | |
| − | + | Various articles [4,5] indicated that oxidative stress increases the activity of certain transcription factors in S. cerevisiae cells. Two transcription factors activated by oxidative stress are Skn7p and Yap1p. These transcription factors have specific binding sites in the promoter areas of genes that respond to oxidative stress. Skn7p binds to OSRE (oxidative stress response element) and Yap1p binds to YRE (Yap1p response element). Some of the best characterized promoter regions that these transcription factors bind to are those for the genes tsa1 (thioredoxin peroxidase) and ccp1 (mitochondrial cytochrome-c peroxidase) [5]. The gene that was reported to have highest response against microcystin was ctt1 (cytosolic catalase T) [2]. We chose to look into these three promoters more deeply, and this led us to the idea of creating a sensor by fusing one of these promoter areas with fluorescent protein production and implementing this in S. cerevisiae. | |
| − | + | </p> | |
| − | + | <br/> | |
| − | + | <p class="justify" style="font-size:19px"> | |
| − | + | This sensor would produce a fluorescent signal when the activation of the transcription factors Skn7p and Yap1p has been induced – in other words, when the S. cerevisiae cells encounter stress. As microcystin is a rather effective stress-generating factor, and as most sources of oxidative stress (such as xenobiotics and heavy metals) are rare in nature [6], we can assume that it is the main stress producing factor found in Finnish lakes. This leads to our hypothesis: fluorescent colour produced by our engineered S. cerevisiae is proportional to microcystin concentration in measured water samples. | |
| − | + | </p> | |
| − | + | <br/> | |
| − | + | <h3> | |
| − | + | Transporter | |
| − | + | </h3> | |
| − | + | <br/> | |
| − | + | <p class="justify" style="font-size:19px"> | |
| − | + | An important prerequisite for the function of the stress promoters is that the toxin must be imported into the cell. Microcystin-LR doesn’t pass the cell membrane without the help of specific transporter proteins. In human liver cells, the membrane bound OATP transporter (organic anion transport peptide) was identified to imports MCs into the cell. OATPs belong to the superfamily of solute carriers and mediate the uptake of endogenous compounds such as hormones or bile salts but also a variety of xenobiotics. [7] In yeast, the QDR2 transporter has a similar function. [8] It has a broad substrate specificity and can transport many mono- and divalent cations as well as a variety or drugs. [9]. According to Valerio et al., the QDR2 transporter from the yeast strain VL3 is the most similar to human OATP out of all S. cerevisiae strains. [2] We also did a BLAST search to confirm this data and come to the conclusion that QDR2 transporter from VL3 corresponded best to the human OATP transporter out of all yeast transporters. Transporters from all other common S. cerevisiae strains differ significantly from these. | |
| − | + | </p> | |
| − | + | <br/> | |
| − | + | <p class="justify" style="font-size:19px"> | |
| − | + | It has been confirmed that the QDR2 transporter from VL3 can import microcystin [2]. We reasoned that by replacing the original QDR2 transporter of a S. cerevisiae lab strain (W303a/SS238) with the VL3 variant of the QDR2 transporter we could enable toxin import in a new strain. The reason for not wanting to work with VL3 was that it’s used and optimized for wine production, not for extensive gene engineering. Another reason for not choosing to engineer VL3 was that we would have had to do auxotrophic mutations for it first as such kind of this strain was not available. However, we planned to use VL3 as a positive control in our experiments to compare how changing the transporter of a lab strain effects observed toxicity effects. This way, we hope to see whether toxicity behaviour corresponding to VL3 can be observed. | |
| − | + | </p> | |
| − | + | <br/> | |
| − | + | <h3> | |
| − | + | Final Product | |
| − | + | </h3> | |
| − | + | <!--<figure style="float: right; text-align:center; padding-left:20px;"> | |
| − | + | <img style="width:80%;" src='img/toxicity.png'><br><figcaption>Figure 1.</figcaption>--> | |
| − | + | <br/> | |
| − | + | <p class="justify" style="font-size:19px"> | |
| − | + | <img src="/wiki/images/d/dd/T--Aalto-Helsinki--toxicity1.png" style="width:50%; float:right;"/> | |
| − | + | For the final product, we will need to have both the VL3 transporter and the stress promoter in yeast cells. The colour of two lake water samples, one supplemented with a defined quantity of engineered S. cerevisiae cells and one with unmodified yeast, can then be compared, and the toxin concentration can be calculated easily from a standard curve utilizing known toxin concentrations. In the final product, we would have a test device containing the measurement and control yeasts. The user could add a small amount of sample water to be measured and analyse it with a mobile app that tells about the toxicity based on the colour difference between the sample and the control. The idea is explained shortly in figure 2. | |
| − | + | </p> | |
| − | + | <br/> | |
| − | + | </div> | |
| − | + | <div class="section" style="font-family: robotolight"> | |
| − | <br/> | + | <h2 style="float:left; margin-left:165px; margin-top: 10px;"> |
| − | + | <i> | |
| − | + | Did you know... | |
| − | + | </i> | |
| − | <i> | + | </h2> |
| − | + | <br/> | |
| + | <br/> | ||
| + | <p style="float:left; margin-left:165px; margin-right:150px;"> | ||
| + | <i> | ||
| + | The oldest known fossils are cyanobacteria from Archaean rocks of western Australia, dated 3.5 billion years old. | ||
| + | </i> | ||
| + | </p> | ||
| + | </div> | ||
| + | <div class="container" style="width: 75%; font-family: robotolight; color: #4d4d33"> | ||
| + | <h1 id="THREE"> | ||
| + | Degradation | ||
| + | </h1> | ||
| + | <br/> | ||
| + | <h3> | ||
| + | Background | ||
| + | </h3> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | The second part of our project, the degradation of the cyanobacterial toxin microcystin (MC), is also engineered in the yeast | ||
| + | <i> | ||
| + | Saccharomyces cerevisiae | ||
| + | </i> | ||
| + | . We want to produce an MC-degrading enzyme called microcystinase (MlrA) which is found naturally in some bacterial species. Our goal is to produce an enzyme capsule which can be added into MC-containing water where it degrades the toxins and makes the water safe to use. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | There is a gene cluster which has been identified to code for enzymes which degrade microcystins. The genes mlrA, mlrB, mlrC and mlrD code for enzymes MlrA, MlrB, MlrC and MlrD respectively. They all have a role in the degradation process with different functions. MlrA, also known as microcystinase, acts first by hydrolysing the bond between microcystin’s amino acids Adda and arginine. This linearizes the toxin, after which MlrB (a serine protease), and MlrC (a metalloprotease) are able to degrade it further to individual amino acids, which are then used in the cell’s metabolism. MlrD’s function is not fully understood at this point, but it may have a role in importing the toxins. (1) | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | Our focus will be on the degradation of MC-LR because it is the most common cyanotoxin and also because the degradation has been characterized for this toxin. The linearization, performed by MlrA, renders the toxin 160 less toxic (2). This should be enough to make for example a bucket of water safe to use as sauna water (check some calculation about this in results somewhere). | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | Since the cyanobacteria release the toxins only as they lyse, we have to take also that into consideration. Based on earlier work by Peking 2014 iGEM team, hen egg lysozyme is good to lyse the tough cell walls of cyanobacteria WE CAN’T USE LYSOZYME. Our final product will contain lysozyme and microcystinase. The lysozyme would first lyse the cyanobacteria and release all the possible MC, and the microcystinase which would then linearize the toxins. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | PICTURE | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | MlrA is a putative membrane protein. It has eight membrane spanning helices and according to our homologous structure model it forms a channel, in middle of which the active site resides. Although MlrA seems to be a membrane protein, there has been studies, where at least some of it is located in the cytosol (3). | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | PICTURE | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | There has been a previous iGEM project about degradation of microcystin (Ranger amongst Enemies, Peking 2014). Their device was engineered in E. coli, which expressed MlrA as a part of their project. The MlrA sequence used was from Sphingomonas, but the exact species was not mentioned. Their BioBrick of the enzyme differed in one amino acid compared to the one we found in GenBank and that we have used in our work. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <h3> | ||
| + | Design | ||
| + | </h3> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | We have three different variants of the enzyme for production in | ||
| + | <i> | ||
| + | S. cerevisiae | ||
| + | </i> | ||
| + | as well as a positive control for production in | ||
| + | <i> | ||
| + | E. coli | ||
| + | </i> | ||
| + | , since it has been already established that heterologous production of MlrA results in active enzyme (3). | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | We will use the MlrA sequence from | ||
| + | <i> | ||
| + | Sphingomonas sp. ACM-396 | ||
| + | </i> | ||
| + | (LINK TO GENBANK), as it has been characterized to degrade MC-LR, and because apparently it was the one used in Peking 2014 team’s work. To make sure that we get the best possible protein yields from expression in yeast and | ||
| + | <i> | ||
| + | E. coli | ||
| + | </i> | ||
| + | , codon optimization was required. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | In mass production the most efficient way to produce proteins is having them secreted into the growing media. This poses a problem as our enzyme is putatively a membrane protein. In the effort of having it secreted, we are using a yeast-specific secretion tag, mating factor α. Mating factor α is also used to localize membrane proteins into the plasma membrane. We will produce four different versions of the enzyme: MlrAY, MlrAYAlpha, MlrAY3Alpha and MlrAE. | ||
| + | </p> | ||
| + | <ul style="font-size:19px"> | ||
| + | <li> | ||
| + | MlrAY, containing no mating factor α, should be soluble in the cytosol. | ||
| + | </li> | ||
| + | <li> | ||
| + | MlrAYAlpha has one secretion tag, which should localize the protein into the plasma membrane. | ||
| + | </li> | ||
| + | <li> | ||
| + | MlrAY3Alpha has a secretion tag with a triple propeptide part, and so is the most likely of the three proteins to be secreted, if at all. | ||
| + | </li> | ||
| + | <li> | ||
| + | MlrAE, used as a positive control by production of the enzyme in E. coli. It has been demonstrated earlier that the enzyme is active in this production system. | ||
| + | </li> | ||
| + | </ul> | ||
| + | <p> | ||
| + | </p> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | For the yeast backbone we are using pRS415-GAL whereas pSVA1 is used as the bacterial backbone. Both are available to us at our lab. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | For purification of the protein, we are putting a 6xHis-tag into the C-terminal of the protein. This should allow for a simple purification using Immobilized Metal Affinity Chromatography (IMAC). If the enzyme localizes in membranes with the secretion tag(s), the best way to purify it while making sure it is still active is to purify the membranes as a whole. This means that the enzyme would be in its natural environment in the purified form. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | In the design of the degradation part of our project we won’t be using BioBricks simply because there aren’t any appropriate ones available/well characterized. Our designs weren’t compatible with the BioBrick standard, and we didn’t want to compromise them in order to have for example a ready BioBrick backbone. | ||
| + | </p> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | PICTURE | ||
| + | </p> | ||
| + | <br/> | ||
| + | <h4> | ||
| + | References | ||
| + | </h4> | ||
| + | <br/> | ||
| + | <p class="justify" style="font-size:19px"> | ||
| + | 1. Bourne et al., 2001, Characterization of a gene cluster… | ||
<br/> | <br/> | ||
| + | 2. Bourne et al., 1996, Enzymatic Pathway for the Bacterial Degradation of the Cyanobacterial Cyclic Peptide Toxin Microcystin LR | ||
<br/> | <br/> | ||
| − | + | 3. Dziga et al., 2012, Heterologous…. of MlrA | |
| − | </ | + | </p> |
| − | </ | + | <br/> |
| + | </div> | ||
</div> | </div> | ||
| − | < | + | <script> |
| − | + | $(window).scroll(function(){ | |
| − | + | $(".title").css("opacity", 1 - $(window).scrollTop() / 500); | |
| − | + | }); | |
| − | + | </script> | |
| − | + | <!-- Bootstrap core JavaScript | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
================================================== --> | ================================================== --> | ||
| − | + | <!-- Placed at the end of the document so the pages load faster --> | |
| − | + | <!--script src="./Tinyone - HTML_files/jquery.min.js"></script> | |
<script src="./Tinyone - HTML_files/bootstrap.min.js"></script> | <script src="./Tinyone - HTML_files/bootstrap.min.js"></script> | ||
<script src="./Tinyone - HTML_files/offcanvas.js"></script> | <script src="./Tinyone - HTML_files/offcanvas.js"></script> | ||
<script src="./Tinyone - HTML_files/function.js"></script--> | <script src="./Tinyone - HTML_files/function.js"></script--> | ||
| + | </div> | ||
</body> | </body> | ||
</html> | </html> | ||
Latest revision as of 04:58, 3 December 2016
PROJECT
Overview
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Suspendisse suscipit arcu libero. Duis vitae cursus elit. Donec enim turpis, aliquam id neque at, faucibus porta odio. Pellentesque habitant morbi tristique senectus et netus et malesuada fames ac turpis egestas. Aliquam et sollicitudin risus. Pellentesque risus magna, accumsan sit amet purus sed, ornare mattis diam. Morbi ac arcu metus. Praesent in massa lectus. Proin fringilla, erat a tempor imperdiet, nunc purus venenatis sem, vel finibus ex lacus a nisi. Morbi nibh augue, ultrices ut lacus id, laoreet aliquam mauris.
Did you know...
Cyanobacteria range in size from 0.5 to 60 micrometers in diameter which represents one of the largest prokaryotic organism.
Detection
Background
When studying the toxicity of the cyanobacterial toxin microcystin-LR in higher eukaryotic cells, we found out that it causes oxidative stress in them. This is due to inhibition of protein phosphatases 1 and 2A (PP1 and PP2A) which then leads to hyperphosphorylation of target proteins. This excessive phosphorylation in turn increases the formation of reactive oxygen species (ROS), resulting in a oxidative stress response. This is presented in the figure 1. [1] Reactive oxygen species have many effects in the cell. For example, catalase production is significantly increased in the presence of ROS. Catalase is an enzyme found in nearly all living organisms that catalyses the decomposition hydrogen peroxide. It is one of the most important enzymes in protecting the cell from reactive oxygen species meaning that its production is activated when there is oxidative stress. We found out that similar toxicity effects can be observed in some lower eukaryotes, such as the yeast Saccharomyces cerevisiae. [2,3] Realizing this made us think of how we could use MC-LR’s natural toxicity effects to sense the toxin concentrations.
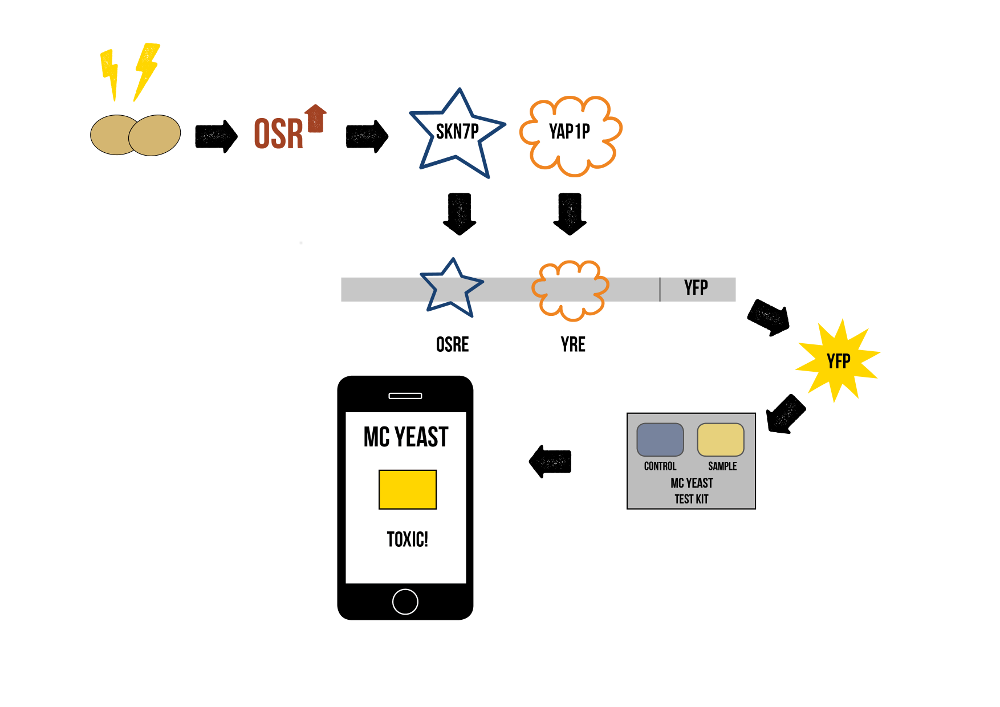
Stress Promoters
Various articles [4,5] indicated that oxidative stress increases the activity of certain transcription factors in S. cerevisiae cells. Two transcription factors activated by oxidative stress are Skn7p and Yap1p. These transcription factors have specific binding sites in the promoter areas of genes that respond to oxidative stress. Skn7p binds to OSRE (oxidative stress response element) and Yap1p binds to YRE (Yap1p response element). Some of the best characterized promoter regions that these transcription factors bind to are those for the genes tsa1 (thioredoxin peroxidase) and ccp1 (mitochondrial cytochrome-c peroxidase) [5]. The gene that was reported to have highest response against microcystin was ctt1 (cytosolic catalase T) [2]. We chose to look into these three promoters more deeply, and this led us to the idea of creating a sensor by fusing one of these promoter areas with fluorescent protein production and implementing this in S. cerevisiae.
This sensor would produce a fluorescent signal when the activation of the transcription factors Skn7p and Yap1p has been induced – in other words, when the S. cerevisiae cells encounter stress. As microcystin is a rather effective stress-generating factor, and as most sources of oxidative stress (such as xenobiotics and heavy metals) are rare in nature [6], we can assume that it is the main stress producing factor found in Finnish lakes. This leads to our hypothesis: fluorescent colour produced by our engineered S. cerevisiae is proportional to microcystin concentration in measured water samples.
Transporter
An important prerequisite for the function of the stress promoters is that the toxin must be imported into the cell. Microcystin-LR doesn’t pass the cell membrane without the help of specific transporter proteins. In human liver cells, the membrane bound OATP transporter (organic anion transport peptide) was identified to imports MCs into the cell. OATPs belong to the superfamily of solute carriers and mediate the uptake of endogenous compounds such as hormones or bile salts but also a variety of xenobiotics. [7] In yeast, the QDR2 transporter has a similar function. [8] It has a broad substrate specificity and can transport many mono- and divalent cations as well as a variety or drugs. [9]. According to Valerio et al., the QDR2 transporter from the yeast strain VL3 is the most similar to human OATP out of all S. cerevisiae strains. [2] We also did a BLAST search to confirm this data and come to the conclusion that QDR2 transporter from VL3 corresponded best to the human OATP transporter out of all yeast transporters. Transporters from all other common S. cerevisiae strains differ significantly from these.
It has been confirmed that the QDR2 transporter from VL3 can import microcystin [2]. We reasoned that by replacing the original QDR2 transporter of a S. cerevisiae lab strain (W303a/SS238) with the VL3 variant of the QDR2 transporter we could enable toxin import in a new strain. The reason for not wanting to work with VL3 was that it’s used and optimized for wine production, not for extensive gene engineering. Another reason for not choosing to engineer VL3 was that we would have had to do auxotrophic mutations for it first as such kind of this strain was not available. However, we planned to use VL3 as a positive control in our experiments to compare how changing the transporter of a lab strain effects observed toxicity effects. This way, we hope to see whether toxicity behaviour corresponding to VL3 can be observed.
Final Product
 For the final product, we will need to have both the VL3 transporter and the stress promoter in yeast cells. The colour of two lake water samples, one supplemented with a defined quantity of engineered S. cerevisiae cells and one with unmodified yeast, can then be compared, and the toxin concentration can be calculated easily from a standard curve utilizing known toxin concentrations. In the final product, we would have a test device containing the measurement and control yeasts. The user could add a small amount of sample water to be measured and analyse it with a mobile app that tells about the toxicity based on the colour difference between the sample and the control. The idea is explained shortly in figure 2.
For the final product, we will need to have both the VL3 transporter and the stress promoter in yeast cells. The colour of two lake water samples, one supplemented with a defined quantity of engineered S. cerevisiae cells and one with unmodified yeast, can then be compared, and the toxin concentration can be calculated easily from a standard curve utilizing known toxin concentrations. In the final product, we would have a test device containing the measurement and control yeasts. The user could add a small amount of sample water to be measured and analyse it with a mobile app that tells about the toxicity based on the colour difference between the sample and the control. The idea is explained shortly in figure 2.
Did you know...
The oldest known fossils are cyanobacteria from Archaean rocks of western Australia, dated 3.5 billion years old.
Degradation
Background
The second part of our project, the degradation of the cyanobacterial toxin microcystin (MC), is also engineered in the yeast Saccharomyces cerevisiae . We want to produce an MC-degrading enzyme called microcystinase (MlrA) which is found naturally in some bacterial species. Our goal is to produce an enzyme capsule which can be added into MC-containing water where it degrades the toxins and makes the water safe to use.
There is a gene cluster which has been identified to code for enzymes which degrade microcystins. The genes mlrA, mlrB, mlrC and mlrD code for enzymes MlrA, MlrB, MlrC and MlrD respectively. They all have a role in the degradation process with different functions. MlrA, also known as microcystinase, acts first by hydrolysing the bond between microcystin’s amino acids Adda and arginine. This linearizes the toxin, after which MlrB (a serine protease), and MlrC (a metalloprotease) are able to degrade it further to individual amino acids, which are then used in the cell’s metabolism. MlrD’s function is not fully understood at this point, but it may have a role in importing the toxins. (1)
Our focus will be on the degradation of MC-LR because it is the most common cyanotoxin and also because the degradation has been characterized for this toxin. The linearization, performed by MlrA, renders the toxin 160 less toxic (2). This should be enough to make for example a bucket of water safe to use as sauna water (check some calculation about this in results somewhere).
Since the cyanobacteria release the toxins only as they lyse, we have to take also that into consideration. Based on earlier work by Peking 2014 iGEM team, hen egg lysozyme is good to lyse the tough cell walls of cyanobacteria WE CAN’T USE LYSOZYME. Our final product will contain lysozyme and microcystinase. The lysozyme would first lyse the cyanobacteria and release all the possible MC, and the microcystinase which would then linearize the toxins.
PICTURE
MlrA is a putative membrane protein. It has eight membrane spanning helices and according to our homologous structure model it forms a channel, in middle of which the active site resides. Although MlrA seems to be a membrane protein, there has been studies, where at least some of it is located in the cytosol (3).
PICTURE
There has been a previous iGEM project about degradation of microcystin (Ranger amongst Enemies, Peking 2014). Their device was engineered in E. coli, which expressed MlrA as a part of their project. The MlrA sequence used was from Sphingomonas, but the exact species was not mentioned. Their BioBrick of the enzyme differed in one amino acid compared to the one we found in GenBank and that we have used in our work.
Design
We have three different variants of the enzyme for production in S. cerevisiae as well as a positive control for production in E. coli , since it has been already established that heterologous production of MlrA results in active enzyme (3).
We will use the MlrA sequence from Sphingomonas sp. ACM-396 (LINK TO GENBANK), as it has been characterized to degrade MC-LR, and because apparently it was the one used in Peking 2014 team’s work. To make sure that we get the best possible protein yields from expression in yeast and E. coli , codon optimization was required.
In mass production the most efficient way to produce proteins is having them secreted into the growing media. This poses a problem as our enzyme is putatively a membrane protein. In the effort of having it secreted, we are using a yeast-specific secretion tag, mating factor α. Mating factor α is also used to localize membrane proteins into the plasma membrane. We will produce four different versions of the enzyme: MlrAY, MlrAYAlpha, MlrAY3Alpha and MlrAE.
- MlrAY, containing no mating factor α, should be soluble in the cytosol.
- MlrAYAlpha has one secretion tag, which should localize the protein into the plasma membrane.
- MlrAY3Alpha has a secretion tag with a triple propeptide part, and so is the most likely of the three proteins to be secreted, if at all.
- MlrAE, used as a positive control by production of the enzyme in E. coli. It has been demonstrated earlier that the enzyme is active in this production system.
For the yeast backbone we are using pRS415-GAL whereas pSVA1 is used as the bacterial backbone. Both are available to us at our lab.
For purification of the protein, we are putting a 6xHis-tag into the C-terminal of the protein. This should allow for a simple purification using Immobilized Metal Affinity Chromatography (IMAC). If the enzyme localizes in membranes with the secretion tag(s), the best way to purify it while making sure it is still active is to purify the membranes as a whole. This means that the enzyme would be in its natural environment in the purified form.
In the design of the degradation part of our project we won’t be using BioBricks simply because there aren’t any appropriate ones available/well characterized. Our designs weren’t compatible with the BioBrick standard, and we didn’t want to compromise them in order to have for example a ready BioBrick backbone.
PICTURE
References
1. Bourne et al., 2001, Characterization of a gene cluster…
2. Bourne et al., 1996, Enzymatic Pathway for the Bacterial Degradation of the Cyanobacterial Cyclic Peptide Toxin Microcystin LR
3. Dziga et al., 2012, Heterologous…. of MlrA