Data recovery
Using a surface slide system (SSS) the dynamics of the plasmid R1 and its permanently derepressed mutant Rldvdl9 in surface cultures of Escherichia coli K12 was examined.
The experiments were carried on E.coli K12 strain J53 hosting the plasmid R1 or it's premanently repressed mutant R1drd19. those two strains were used as donor cells. The recipient used is E.coli CH50.
Here is a summary of their result on the effect of initial cell density on bacterial growth and plasmid transfer in surface and liquide culture.
| Surface mating JS3(RI) x CSHSO |
Surface mating J53(Rldrd19) x CSHSO
|
| Initial |
Recipient |
Donor |
Transconjugant |
Total |
Initial |
Recipient |
Donor |
Transconjugant |
Total
|
| 10^7 |
3,8*10^8 |
1,5*10^8 |
2,1*10^5 |
5,2*10^8 |
10^7 |
6,6*10^7 |
1,7*10^8 |
9,4*10^7 |
3,3*10^8
|
| 10^6 |
3,1*10^8 |
1,8*10^8 |
2,1*10^5 |
4,8*10^8 |
10^6 |
1,4*10^8 |
2,5*10^8 |
4,4*10^7 |
4,4*10^8
|
| 10^5 |
2,2*10^8 |
1,8*10^8 |
6,5*10^4 |
4,1*10^8 |
10^5 |
1,4*10^8 |
1,9*10^8 |
8,0*10^6 |
3,4*10^8
|
| 10^4 |
2,7*10^8 |
1,9*10^8 |
3,0*10^4 |
4,6*10^8 |
10^4 |
1,6*10^8 |
2,3*10^8 |
1,8*10^6 |
3,9*10^8
|
| 10^3 |
2,7*10^8 |
1,9*10^8 |
3,0*10^4 |
4,6*10^8 |
10^3 |
2,3*10^8 |
2,6*10^8 |
4,7*10^5 |
4,9*10^8
|
| 10^2 |
2,9*10^8 |
2,2*10^8 |
1,6*10^4 |
5,0*10^8 |
10^2 |
2,4*10^8 |
2,2*10^8 |
4,1*10^4 |
4,7*10^8
|
| Liquid mating JS3(RI) x CSHSO |
Liquid mating JS3(Rldrd19) x CSHSO
|
| Initial |
Recipient |
Donor |
Transconjugant |
Total |
Initial |
Recipient |
Donor |
Transconjugant |
Total
|
| 10^7 |
2,2*10^8 |
9,8*10^7 |
4,4*10^3 |
3,1*10^8 |
10^7 |
1,2*10^8 |
6,8*10^7 |
7,4*10^7 |
2,6*10^8
|
| 10^6 |
1,7*10^8 |
8,5*10^7 |
5,9*10^3 |
2,6*10^8 |
10^6 |
1,7*10^8 |
3,7*10^7 |
8,4*10^7 |
2,9*10^8
|
| 10^5 |
2,0*10^8 |
6,9*10^7 |
7,9*10^3 |
2,7*10^8 |
10^5 |
1,7*10^8 |
2,7*10^7 |
7,4*10^7 |
2,7*10^8
|
| 10^4 |
2,7*10^8 |
6,1*10^7 |
1,6*10^4 |
2,7*10^8 |
10^4 |
1,9*10^8 |
1,8*10^7 |
6,6*10^7 |
2,7*10^8
|
| 10^3 |
2,9*10^8 |
4,9*10^7 |
1,9*10^4 |
2,9*10^8 |
10^3 |
2,3*10^8 |
7,4*10^6 |
4,7*10^7 |
2,8*10^8
|
| 10^2 |
4,1*10^8 |
1,0*10^7 |
6,0*10^3 |
4,1*10^8 |
10^2 |
2,6*10^8 |
3,7*10^6 |
1,2*10^7 |
2,8*10^8
|
They have made the following curves to show their results.
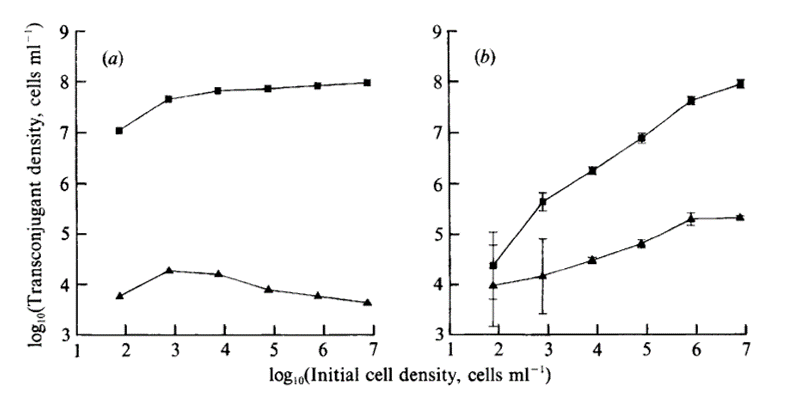
Dynamics of surface and liquid matings for the plasmids R1 and Rldrdl9 for different initial densities. Transconjugant production (cells per ml medium at stationary phase) in (a) liquid cultures and (b) surface cultures is shown. The plotted points for the liquid cultures represent means; those for the surface cultures represent means 2 x SE. Squares, Matings with Rldrdl9; Triangles, matings with R1.
Their conclusion on the plasmid transfer rate are:
On SSS, when colony density is high, many colonies meet, resulting in the original donors being responsible for most transconjugant production. At low colony densities, few colonies meet, and transitorily derepressed transconjugants are the primary donors. This hypothesis is supported by the observation that transconjugant yields for Rldrdl9 and R1 were similar at the lowest colony density, suggesting that in both cases the primary donors were derepressed.
On liquid batch cultures, transitory derepression does not play a significant role, since during the growth period the derepressed transconjugants are a very small fraction of the randomly distributed donor populations (Levin et al., 1979).

{kind=link}