Violacein is a fluorescent reporter with anticancer activity (Carvalho et. al, 2006)
that has been used in several other iGEM projects (Cambridge 2009, Slovenia 2010,
Johns Hopkins 2011, UCSF 2012). Although it would be a good pigment candidate for
our project, it has a complex synthetic pathway requiring five specialized enzymes
and oxygen (Fig. 2). It also presents multiple off-path reactions that can reduce
the efficiency of the pathway. Before building constructs to use for violacein
production, we needed to find a way to determine which promoters to use for
the five genes involved in the pathway. Although there are studies focused on
the optimization of the production of violacein (Lee et. al, 2013), none of
the studies give a biochemical model of the rates of the reactions that take
place in the bacteria.
Create a biochemical model of the violacein production based on the synthetic
pathway and violacein production data from bacteria with different promoters for
each of the five genes involved in the pathway.
1. Modeling Promoter Strength
Because a major goal of the model is to predict the effects of the selection of
promoters on the final production of violacein, we decided to find a way to
characterize promoters first. To simplify the computation, we used the promoter
strength as a single standard to characterize the promoters. Moreover, we assumed
the degradation rate of proteins only depends on the growth rate of E.coli. Then,
every enzyme has the same degradation rate. The bacteriophage T7 promoter has
been widely used for protein expression and purification (Jones et al., 2013),
so we used data of five mutant T7 promoters to create a proof-of-concept model.
If this model was functional, we could implement the same modeling technique to
the promoters we were working with.The five mutant T7 promoters have distinct
promoter strength over time after induction. The experimental data from the
literature are shown in the figure below (Jones et al., 2013).
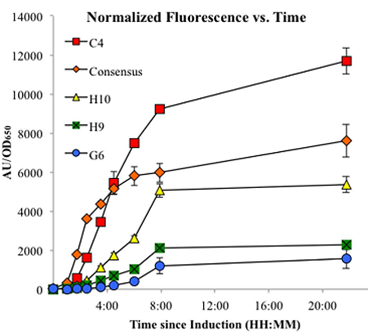
The first step of our model is to describe the rate of change of enzymes based on promoter strength. Here we assumed that the enzyme production rate is directly proportional to strength of the promoter. Therefore, we were able to use a mass-action kinetics equation of promoters to describe the enzyme concentration. The equation is shown below:

In this equation, Ai is the concentration of enzyme i, ki is the production rate of each enzyme i, kd is the degradation rate of all enzymes, and t is time. By solving this equation, we derived the equation of enzyme concentration against time.

Since we assumed that the promoter strength is proportional to the promoter concentration, we would use the equation to fit our data using least squares method (Fig. 1).
 Figure 1.
Figure 1. Linear regressions fitted to normalized fluorescence vs time. The circles represent data from Jones et al., 2013. The solid lines are our regression lines. The colors indicate with which promoters the circles and lines correspond.
In general, the regression lines are able to capture the change of strength of each enzyme over time. In this way, the parameters are determined. The table below lists the parameter values.
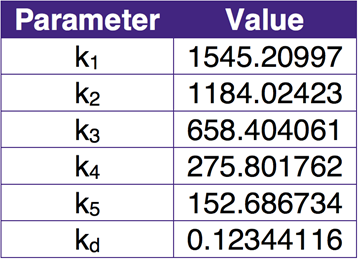
In the table, ki (i = 1,2,3,4,5) are the production rate coefficients of promoter I (i = 1,2,3,4,5), and kd is the degradation rate coefficient of all promoters.
2. Modeling the Steady-state Violacein Yield
After we finished the regression model of each promoter, we created a second model to describe the violacein biosynthetic pathway. The pathway (Fig. 2) involves five enzyme-catalyzed reactions and one non-enzymatic reaction (Lee et al, 2013).
 Figure 2.
Figure 2. Violacein synthetic pathway. The purple arrows highlight the five enzymatic and one non-enzymatic steps of violacein production from two molecules of tryptophan. The five enzymes are indicated by bolding (VioA, VioB, etc.).
The model was developed as three major parts. A pseudocode of this model is provided here.
Define ODE System
- Calculate the production and degradation rate of each molecule in the pathway from the concentration of reagents and parameters.
- Obtain the rate of change of each molecule based on the production and degradation rates.
Solve the System of Nonlinear Equations at Steady State
- Solve the system of nonlinear equations at steady state starting at an initial guess X0.
- Use the result as a new initial guess; repeat the numerical method to solve the system of equations again.
- Calculate the relative error of each chemical in the new result.
- If the maximum error is smaller than 0.0001%, output violacein concentration at steady state as the final result.
Optimize Parameters to Fit Experimental Data
- Set the initial guess of the parameters.
- Load the data from literature, which include the choice of promoter for each gene and the corresponding violacein yield determined experimentally.
- For each promoter selection scenario, pass each promoter numbers and the temporary parameters to the steady-state model.
- Obtain the violacein yield predicted by the steady-state model for each promoter selection scenario.
- Compute the residual sum of squares (RSS) of between the predicted violacein yields and the violacein yields given by experiment.
- Determine the optimal parameters by minimizing the RSS (least square method).
 Figure 3.
Figure 3. VIolacein yield with different promoter combinations. This graph compares the violacein found for various promoter combinations determined by Jones et al., 2013 (shown in blue) with the violacein concentrations that our model predicted for the same promoter combinations.
Discussion
References
- Carvalho, D. D., Costa, F. T. M., Duran, N., & Haun, M. (2006). Cytotoxic activity of violacein in human colon cancer cells. Toxicology in Vitro, 20(8), 1514–1521. http://doi.org/http://dx.doi.org/10.1016/j.tiv.2006.06.007
- Jones, J. A., Vernacchio, V. R., Lachance, D. M., Lebovich, M., Fu, L., Shirke, A. N., … Koffas, M. A. G. (2015). ePathOptimize: A Combinatorial Approach for Transcriptional Balancing of Metabolic Pathways. Scientific Reports, 5, 11301. http://doi.org/10.1038/srep11301
- Lee, M. E., Aswani, A., Han, A. S., Tomlin, C. J., & Dueber, J. E. (2013). Expression-level optimization of a multi-enzyme pathway in the absence of a high-throughput assay. Nucleic Acids Research, 41(22), 10668–10678. http://doi.org/10.1093/nar/gkt809