Difference between revisions of "Team:Tianjin/Experiment/CFPS"
| Line 166: | Line 166: | ||
<div class="col-md-8"> | <div class="col-md-8"> | ||
| − | <img src="https://static.igem.org/mediawiki/2016/ | + | <img src="https://static.igem.org/mediawiki/2016/c/ca/T--Tianjin--cf-e3.png" alt="desktop"> |
<p style="font-size:15px"> | <p style="font-size:15px"> | ||
<br/> | <br/> | ||
Revision as of 13:15, 11 October 2016
Experiment of CFPS
Overview
The ability to produce a functional protein in the test tube, rather than in cells, is the essence of cell-free protein synthesis (CFPS)[1]. .

Fig.1. The basic process of CFPS system[2]
The lack of high-throughput approaches for expression and screening of large enzyme libraries remains a major bottleneck for current enzyme engineering efforts. To address this need, some researchers[2] have developed a high-throughput, fluorescence-based approach for rapid one-pot, microscale expression, and screening of different kinds of enzymes. To go further, we try to make our effert to achieve integration of cell-free protein expression with activity screening of enzymes( site-directed mutations of PETase). .

Fig.2.One-pot approach for integrated expression and activity screening of enzymes
Background
At its core, synthetic biology is inspired by the power and diversity of the living world. It is an endeavor predicated on the idea that we can learn to more reliably and rapidly engineer biological function for compelling applications in medicine, biotechnology, and green chemistry. What is unique to synthetic biology is the application of an engineering-driven approach to accelerate the design-build-test loops required for reprogramming existing, and constructing new, biological systems. This is needed because the current approach to engineering cells is often extremely laborious, costly, and difficult. So far, efforts in synthetic biology have largely focused on the construction and implementation of genetic circuits, biological modules (compilations of biological ‘‘parts’’ that perform defined functions, e.g., ribosome binding sites, promoters, and genes), and synthetic pathways into genetically reprogrammed organisms. However, a major challenge exists because of our incomplete knowledge of how life works, the daunting complexity of cells, the unintended interference between native and synthetic parts, and (unlike typical engineered systems) the fact that cells evolve, have noise, and have their own agenda such as growth and adaptation. The guiding question, therefore, is how do we develop a new way of engineering in the face of these unique and complex features of biology? In other words, what is the suite of foundational technologies required for taming the complexity of living systems? One foundational technology that has emerged is the ability to construct new programs that carry out user-defined functions from ‘‘parts’’ of DNA (e.g., biological switches or oscillators). Biological computer-aided design (BioCAD) tools, directed evolution, and cell-free systems will also each have a role. Although in vivo synthetic biology projects are dominant in the field, we focus this review on the role of cell-free synthetic biology. Without the need to support ancillary processes required for cell viability and growth, cell-free systems offer a powerful platform for accelerating the optimization of synthetic pathways and for not only harnessing but also expanding the chemistry of life.
The power of cell-free systems was first appreciated over a hundred years ago. In 1897, Eduard Buchner used yeast extract to convert sugar to ethanol and carbon dioxide, for which he won the Nobel Prize (1907 Chemistry). Since then, cell-free systems have continued to play an important role in our understanding of fundamental biology. For example, Nirenberg’s discovery of the genetic code, for which he received the Nobel Prize in 1968, shed light into how protein synthesis works. Cell-free systems have since continued to unravel other fundamental discoveries such as the understanding of eukaryotic translation. Beyond using cell-free systems for biochemical analysis, the successful recapitulation of biological function in vitro has inspired attempts to use cell-free systems for product synthesis. Indeed, there are more than 30 years of successful industrial history with biotransformations in vitro. For the most part, in vitro biocatalytic transformation has focused on single transformations, such as enabling the production of specific chirality. In another example, single-enzyme cell-free systems, such as cell-free DNA replication (the polymerase chain reaction (PCR)) are ubiquitous to molecular biology labs and have revolutionized modern biological research. PCR in combination with other common molecular tools such as DNA restriction digest and ligation reactions allow us to assemble DNA sequences at will. In other approaches, crude extract cell-free systems (CECFs) and synthetic enzymatic pathways (SEPs) have practical applications in protein and peptide synthesis and evolution, small molecule production, cellular/metabolic pathway investigation, and non-natural product synthesis.
As one of the most common examples of CECFs, cell-free protein synthesis (CFPS) provides a useful starting point for examining the utility of cell-free systems. To produce target proteins of interest, CFPS systems utilize ensembles of biocatalysts that carry out translation from crude extracts derived from bacterial, plant, or animal cells. Upon incubation with essential amino acids and energy substrates, the ensemble of activated catalysts within the cell lysate acts as a chemical factory to polymerize amino acids into polypeptides. Although any organism can be used to provide a source of crude lysate, the most productive CECFs are derived from Escherichia coli extract.[3]
The translation machinery is the engine of life. In E. coli CFPS the translation machinery is typically about 20-fold more dilute than in thecell, decreasing the rates of initiation, elongation and protein accumulation . As well, the average distance between two adjacent ribosomes on a single mRNA strand increases and polysomes are less likely to form. Despite these differences, CFPS can benefit from the relative slower synthesis rate and the distance between ribosomes by allowing nascent polypeptide chain more time and space to form desirable intra-peptide chain contacts, while decreasing the probability for undesirable, non-specific inter-peptide chain contacts, thereby increasing the probability of proper folding and decreasing the probability of aggregation.
Cell-free protein synthesis is a widely used method in molecular biology. Production of proteins using cell-free protein synthesis usually takes a few hours, in contrast to production of proteins in cells, which typically takes days to weeks. In fact, even first-time users can often obtain newly synthesized proteins in one day using a commercial system.
The diversity of the cell-free systems allows in vitro synthesis of a wide range of proteins for a variety of downstream applications, such as screeening of enzymes activities. In the post-genomic era, cell-free protein synthesis has rapidly become the preferred approach for high-throughput functional and structural studies of proteins and a versatile tool for in vitro protein evolution and synthetic biology.[4]
Experiment Design
Basically, we utilized the cell-free system to express the enzymes which had been modified in 22 different sites. Besides, we added a fluorescet protein, CFP, before the enzyme. And there is a flexible linker, GGGGSGGGGS , between them. So that we could detect the expression of enzymes by detecting expression of the fluorescent protein with a fluorescence readout instrument, for example, a microplate reader. We conceived that with this method we could acquire the best modifications by screening them in a high-throughput way. Then we used the proteins we got to degrade PET.
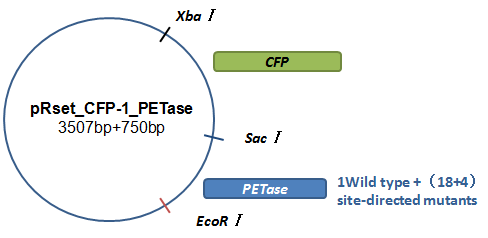
Fig.3.The expression vector in CFPS system
How to characterize the degradation velocity is the main problem in our scheme. We analyzed the experiment consequences in two ways. For the first one, we rendered the enzymes degrade pNPa, a general substituent for the detection of PET. Then we measured the absorbance of pNP in the optical density of 400 nanometers, which is the degrading product of pNPa. For the second one, we detected the absorbance of MHET in the optical density of 260 nanometers, which is the product in the first step of PET degradation.

Fig.4.Steps for integrated expression and activity screening of enzymes
Reference
[1]Gabriel Rosenblum and Barry S. Cooperman. Engine out of the Chassis: Cell-Free Protein Synthesis and its Uses. FEBS Lett. 2014 January 21; 588(2): 261–268. doi:10.1016/j.febslet.2013.10.016.
[2]Aarthi Chandrasekaran and Anup K. Singh. One-Pot, Microscale Cell-Free Enzyme Expression and Screening. DOI 10.1007/978-1-62703-782-2 Springer New York Heidelberg Dordrecht London.
[3] C. Eric Hodgman, Michael C. Jewett. Cell-free synthetic biology: Thinking outside the cell. Metabolic Engineering 14 (2012) 261–269. doi:10.1016/j.ymben.2011.09.002
[4] Shaorong Chong. Overview of Cell-Free Protein Synthesis: Historic Landmarks, Commercial
Systems, and Expanding Applications. Current Protocols in Molecular Biology 16.30.1-16.30.11, October 2014
DOI: 10.1002/0471142727.mb1630s108